ISSN : ISSN: 2576-1412
Journal of Applied Microbiology and Biochemistry
Role of Microsymbionts in Plant Microbe Symbiosis
Mohsin Tariq1,2, Sohail Hameed2, Hikmat Ullah Khan1,3, Muhammad Ikram Munir1, Faryha Nushin1, Saliha Jamil1, Samia Nawaz1 and Marriam Zafar4
1Government College University Faisalabad, Pakistan
2National Institute for Biotechnology and Genetic Engineering (NIBGE), Faisalabad, Pakistan
3Gomal University, Dera Ismail Khan, Pakistan
4Centre of Agricultural Biochemistry and Biotechnology (CABB), University of Agriculture Faisalabad, Pakistan
- *Corresponding Author:
- Mohsin Tariq
Government College University Faisalabad, Pakistan
Tel: 0092-333-6601612
E-Mail: mohsintariq@gcuf.edu.pk; mruaf@hotmail.com
Received Date: March 27, 2017; Accepted Date: April 06, 2017; Published Date: April 13, 2017
Citation: Tariq M, Hameed S, Khan hu , et al. (2017) Role of Microsymbionts in Plant Microbe Symbiosis. J Appl Microbiol Biochem. Vol. 1 No. 2:6
Abstract
Symbiosis refers to the long-term relationship of two unlike species. Microbes symbiotically associate with plant for several reasons, most importantly, to utilize carbon compounds from the root exudates as nutrients. Most of the microsymbionts acknowledge plant provided nutrients via benefitting plant by converting atmospheric nitrogen to the plant usable form of nitrogen, solubilizing unavailable form of phosphate and zinc into available form, providing phytohormones for better plant health, and protecting plant from the pathogens and abiotic stresses. Considering the beneficial properties of plant associating microsymbionts, it could be used as biofertilizers at commercial scale. Biofertilizer is an ecofriendly solution to minimize the hazards of chemical fertilizers. Microsymbionts exert their positive impact on plant once they appropriately colonize rhizosphere or invade plant root or nodules. This review indicates that how microsymbionts colonize plant and contribute to the plant health and crop production.
Keywords
Nodulation; Nitrogen fixation; Phytohormones production; Phosphate solubilization; Biological control
Introduction
Plant microbe symbiosis has significant importance considering its crucial role in nutrient cycling. Microsymbionts of this relationship, microbes, display several important plant growth promoting features. Specifically, most of the rhizosphereassociated microbes have the potential for plant growth stimulation and managing soil health by establishing a symbiotic relationship with the compatible plant. Most extensively studied group of plant beneficial microsymbionts is known as plant growth promoting rhizobacteria (PGPR), which generally colonize the plant root surface and soil adhered to root. Some opportunistic PGPR can enter the root interior, pass through the root cortex and proliferate as endophytes in nodules, tubers, stem, leaves and other organs of plant [1-3]. These bacteria either on the root surface or the plant interior, selectively adapt to the environment and establish a symbiotic relationship with plant [4]. Plant secrete a substantial amount of the carbon fixed, 5-21%, mainly as root exudates, which is the chief source of attraction and nutrition for microbe and initiate a symbiotic relationship with microsymbionts. Microsymbionts in response promote plant growth by making the nutrients available, phytohormones production and controlling plant diseases [5].
Nodulating microsymbionts
Root-nodulating and nitrogen fixing bacteria are collectively termed as ‘rhizobia’ and traditionally belong to one of the six genera; Allorhizobium, Azorhizobium, Bradyrhizobium, Ensifer (formerly Sinorhizobium), Mesorhizobium and Rhizobium [6]. Molecular data have shown that rhizobial bacteria actually constitute a polyphyletic assemblage of alpha, beta and gamma proteobacteria, which collectively form the order Rhizobiales [7-9]. The rhizobia-legume symbiosis exhibits great host specificity, but so far complete information of host specificity is not available. Nodulation factor produced by rhizobia are the key components for the host specificity. A major breakthrough in rhizobia- legume symbioses research was the identification of Nod factor structure from Sinorhizobium meliloti [10]. Currently, Nod factor populations and structure produced by rhizobial strains varies depending upon the type of rhizobia and its geographical location. Nod factors are lipo-chitooligosaccharides, which are considered as molecular keys for the entry of microsymbionts into the legume host. Nod factors generally consist of a backbone of three to five rings of β-1,4-linked N-acetylglucosamine (G1cNAc), which is decorated at nonreducing-terminal with N-methyl, O-acetyl, N-acylated or O-carbamoyl groups and reducing-terminal carry acetyl, L-fucosyl, sulfate ester, 4-O-Ac-fucosyl or 2-O-Me-fucosyl (Figure 1) [11].

Figure 1: General structure of Nod factor.
Interestingly, some Nod factors have exceptional length and type of G1cNAc backbone and diversity in its terminal’s substitutions. Nod factors isolated from M. loci NZP2213 consist of a dimeric chito-oligosaccharidic backbone [12], whereas a pentameric Nod factor whose middle G1cNAc residue is replaced by a glucosyl group is explored from Sinorhizobium fredii [13]. Rhizobium sp. GRH2 synthesize Nod factors consists of six G1cNAc residues [14]. Variable backbone length and substitutions at the reducing end also determine Nod factor host specificity [11]. Perception of Nod factor on plant root surface elicits a series of responses including root hair deformation, membrane potential depolarization, changes in ion fluxes, intracellular and extracellular alkalinization, induction of nodulin gene (ENOD) expression and nodule primordial development, which finally results in the mature nitrogen-fixing nodules formation [15].
Associative microsymbionts
Associative microsymbionts mostly live in the phyllosphere, rhizosphere and rhizoplane or sometimes reside inside the plant tissue and organ. Rhizosphere and rhizoplane associated bacteria are termed as rhizobacteria, whereas those strictly found inside the plant are known as endophytic bacteria. The term rhizoplane refers to the root surface, whereas, the rhizosphere is the immediate thin zone of soil surrounding the root. Legume root nodules also accommodate various non-nodulating plant growth promoting endophytic bacteria, which influence the survival of rhizobia and impact the nodulation and grain yield of crop [16]. The influence associative bacteria may be passive, but most often they promote plant growth. Phyllosphere refers to the aerial part of the plant that also carries microorganisms. Bacteria are the most extensively studied microorganisms in this part of plant. Rhizosphere and rhizoplane associating bacteria generally resides in the soil and move towards the plant as it produces root exudates. Microsymbionts attach themselves with to the root surface via cell surface polysaccharides or cell structures including fimbriae and flagella. Bacteria, cyanobacteria, fungi and algae reside in and around the root zone but bacteria is the most abundant [17]. The associative microsymbionts establish a relationship with plant for nutrient and shelter and in return promote plant growth mainly by fixing nitrogen, indole acetic acid production, phosphate solubilization, biocontrol of plant diseases and by many other mechanisms, which are being explored.
Characteristics of microsymbionts
Biological nitrogen fixation: Nitrogen is one of the most important plant nutrient applied exogenously to fulfill the world’s food demand. Extensive application of chemical fertilizers has resulted serious hazards to the environment and human health. Nitrogen fixed by bacteria is environment friendly, replenishes nitrogen content of the biosphere and important for sustainable agriculture. Biological nitrogen fixation “the conversion of atmospheric nitrogen into ammonia” by the plant microsymbionts is a process of tremendous importance for food security and environment. These nitrogen-fixing bacteria are categorized as diazotrophs. Some bacteria have the ability to fix nitrogen as free N fixer, while other associate plant and develop nodules on the plant roots. Inside the nodules, oxygen free environment is created with the help an indigenous protein leghaemoglobin. Anaerobic bacteria, rhizobia, fix atmospheric nitrogen in this compartment. Nitrogen fixed by biological methods is eco-friendly and does not leach down or evaporates as nitrous oxide to pollute environment. Nitrogen fixation system and machinery does not exist in eukaryotes. The ability to fix nitrogen is distributed amongst a few microorganisms. Nitrogen fixation ability is found in most Eubacterial phylogenetic groups, including Actinomycetes, Cyanobacteria, Proteobacteria, green sulphur bacteria and Firmibacteria. Only a very few Archaea have the ability to fix nitrogen [18].
The enzyme responsible for the conversion of atmospheric dinitrogen to ammonia is known as Nitrogenase. It is a complex metallo-enzyme consisting of two components, smaller dimeric component functions as an ATP-dependent electron donor called iron (Fe) protein, and larger hetero-tetrameric component contains catalytic site of the enzyme called molybdenum–iron (MoFe) protein. Both of the nitrogenase component proteins are extremely oxygen sensitive [19]. The overall stoichiometry of dinitrogen reduction in optimal conditions is as under;
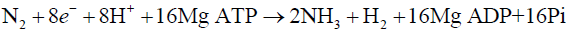
Indole acetic acid production: Indole-3-acetic acid (IAA) is one of the major hormones belongs to the auxins family. It is an important hormone that plays a significant role in many aspects of bacterial life cycle including virulence induction, cell cycle regulation and acid resistance. It also controls many important physiological processes including cell enlargement, cell division and tissue differentiation in responses to light and gravity. By modifying certain conditions like, increase in osmotic content of cell, increase in permeability of water into cell, decrease in wall pressure, an increase in cell wall synthesis and protein synthesis, IAA can stimulate cell elongation. IAA can also inhibit abscission of leaves and induce flowering as well as fruiting [20]. IAA is important for young seedlings as it enhances their ability to anchor themselves to the soil and to obtain maximum water and nutrients from their surrounding environment, so they may able to survive It was later found that various microorganisms including bacteria, fungi, and algae are capable of producing physiologically active quantities of auxins, which may exert pronounced effects on plant growth and establishment [21]. IAA can be produced by many rhizospheric bacteria that inhabitate plants. Due to the rich supplies of substrates exuded from the roots of plants, these rhizospheric bacteria synthesize and release IAA as secondary metabolites that directly promote plant growth. Tryptophan is generally considered as a precursor of IAA. The precursor of auxin as present in root exudate is usually synthesized from the exudate amino acid called tryptophan. IAA is the common product of L-tryptophan metabolism produced by several microorganisms that includes PGPR. The tryptophan concentration in exudate differs strongly among plants. Production of IAA by microbial isolates varies greatly among different species and strains of the same species and is influenced by culture condition, growth stage, and availability of the substrates [22]. The IAA produced by bacteria colonizing the rhizosphere of plants is proposed to act in conjunction with endogenous IAA in plant to stimulate cell proliferation, elongation, and enhancement of host’s uptake of minerals and nutrients from the soil [23]. In addition, IAA serves as a regulating agent for microbial cell differentiation; for example, it stimulates spore germination and mycelial elongation in a streptomycetes, and induces adhesion and ﬠlamentation in Saccharomyces cerevisiae [24,25].
Phosphate solubilization: Phosphorus (P) is major essential macronutrients for plant growth activity. One of the major constituent to mobilize energy in the plant and participate in functional integrity of nucleic acids. P is instituted in the soil in two basic formats, organic and the inorganic. The larger fraction of P prevailed in the soil in insoluble reservoirs include hydroxyapatite, and apatite. The deranged soluble P in soil matrix blemish the plant life activity. To recuperate the plant life activity towards betterment soil need to be provided with avenue of P most readily endorsed by plant. For the flourishing trade of soluble dish of phosphate to the soil is made available by chemical fertilizers, therefore P is applied to soil in the form of phosphatic fertilizers. During the aisle of chemical fertilizers in the soil and over time majority of the fertilizers embedded in the insoluble pool of P in the soil matrix. These insoluble preserve include tricalcium phosphate, aluminum phosphate, and dicalcium phosphate.
Revitalizing plant with a substantial fraction of the inorganic Phosphorus unfortunately is claimed by immobilized pool of P in the soil matrix [26]. Soil microbes house a noble trait that can recuperate the immobilized P into Phosphate. The soil microorganism maneuver through exhaustive solubilization and mineralization, read the soil for inorganic P and ultimately regenerate in soluble form [27]. Cofactor pyrrolquinoline quinone (PQQ) is found to be involved in P solubilization. The current perspective is to rescue soluble P loss, and score a quality production. This refreshing attribute of phosphate solubilization has increased the participation of agronomists in deploying the phosphate solubilizing bacteria as inoculums [28]. The avarice of bacteria to solubilize phosphate has positive influence on Plant life cycle. This compliance of bacteria in the agro-ecosystem has poured a key advantage over the application of chemical fertilizers due to effectiveness, and low cost [29].
Biological control: Annual crop losses due to plant diseases are responsible for a total value of more than 200 billion dollars. Resistant plants and chemicals are often used to control plant disease. Resistance does not exist against all diseases and the breeding of resistant plants takes many years. Moreover, acceptance of genetically engineered resistance is still a sensitive issue in the European Union as so many risk factors are involved. Consumers and supermarket chains are negatively perceived the use of agrochemicals. Unprecedented use of agrochemicals results various negative effects including pathogen resistance and elimination of non-target organism. Most of the agrochemicals applied do not reach the plant. Moreover, consumer desire for pesticide free food and the increasing price of pesticides, especially in poor areas of the world, has led to a search for alternative methods to protect crops. Additionally, Chemical solutions are hardly few, inefficient or nonexistent for many particular diseases. It is ever more prohibited by governmental policies [30].
Biological control, reduction and elimination of pathogen using living organism, is an attractive and environment-friendly solution to combat plant disease and reduce the use of chemical pesticides. Microorganisms are natural competitor of the pathogen. Microbes produce secondary metabolites, locally, onto or near the plant surface to eliminate or inhibit the growth of pathogen [31]. These secondary metabolites are biodegradable and do not cause environmental pollution, compared to the agrochemicals that are designed to resist degradation by microbes. Rhizobacteria are usually studied for antagonism against pathogenic bacteria or fungi. Some rhizobacteria perform antagonistic activity against insects and weeds also [32-34].
Enzyme 1-aminocyclopropane-1- carboxylate (ACC) deaminase present in PGPR functions to promote plant growth by reducing the quantity of ethylene in plant. ACC deaminase producing bacteria take up the ethylene precursor ACC and transform it into 2-oxobutanoate and NH3. There are many forms of stress that are relieved by ACC deaminase producers, like resistance to stress from polyaromatic hydrocarbons, from heavy metals such as Ca2+ and Ni2+, from salt and draught and effects of phytopathogenic bacteria [34,35].
Conclusion and Future Aspects
Plant symbiotic microbes could be isolated from the soil, rhizosphere, plant interior, nodules and could be used as biofertilizers. Microsymbionts improves plant health and crop production as well as enhances soil fertility. One of the important and emerging characteristics of microsymbiont is their antagonistic nature that prevents the plant from various diseases. Researchers are working on biofertilizers since many decades, generated fruitful results and commercially viable products. Microbial inoculants are stable for long term in suitable field environment and contribute major role in sustainable agriculture. There is continuous need to explore new and novel microsymbionts as environmental changes occur over time and existing microbiota may lose it potential.
References
- Kloepper JW, Rodriguez UR, Zehnder GW, Murphy JF, Sikora E, et al. (1999) Plant root bacterial interactions in biological control of soil borne diseases and potential extension to systemic and foliar diseases. Austral Plant Pathol 28: 21-26.
- Gray EJ, Smith DL (2005) Intracellular and extracellular PGPR commonalities and distinctions in the plant bacterium signaling processes. Soil BiolBiochem 37: 395-412.
- Compant S, Reiter B, Sessitsch A, Nowak J, Clement C, et al. (2005) Endophytic colonization of Vitisvinifera L. by a plant growth promoting bacterium Burkholderiasp strain PsJN. Appl Environ Microbiol 71: 1685-1693.
- Compant S, Duffy B, Nowak J, Clement C, Barka EA, et al. (2005) Use of plant growth promoting bacteria for biocontrol of plant diseases principles mechanisms of action and future prospects. Appl Environ Microbiol 71: 4951-4959.
- Lugtenberg B, Kamilova F (2009) plant growth promoting rhizobacteria. Annu Rev Microbiol 63: 541-556.
- Sawada H, Kuykendall LD, Young JM (2003) Changing concepts in the systematics of bacterial nitrogen-fixing legume symbionts. J Gen ApplMicrobiol 49: 155-179.
- Benhizia Y, Benhizia H, Benguedouar A, Muresu R, Giacomini A, et al. (2004) Gamma-proteobacteria can nodulate legumes of the genus Hedysarum. SystApplMicrobiol 27: 462-468.
- Angus A, Hirsch AM (2010) Insights into the history of the legume beta-proteobacterial Symbiosis. MolEcol 19: 28-30.
- Bontemps C, Elliott GN, Simon MF (2010) Burkholderia species are ancient symbionts of legumes. MolEcol 19: 44-52.
- Lerouge P, Roche P, Faucher C, Maillet F, Truchet G, et al. (1990) Symbiotic host specificity of Rhizobium meliloti is determined by a sulphated and acylated glucose oligosaccharide signal. Nature 344: 781-784.
- D'Haeze W, Holsters M (2002) Nod factor structures responses and perception during initiation of nodule development. GlycoBiol 12: 79-105.
- Olsthoorn MMA, López Lara IM, Petersen BO, Bock K, Haverkamp J, et al. (1998) Novel branched Nod factor structure results from α-(1→3) fucosyltransferase activity: the major lipo-chitin oligosaccharides from Mesorhizobium loti strain NZP2213 bear an α-(1→3) fucosyl substituent on a nonterminal backbone residue. Biochem 37: 9024-9032.
- BecFerte MP, Krishnan HB, Savagnac A, Pueppke SG, Prome JC, et al. (1996) Rhizobium fredii synthesizes an array of lipo oligosaccharides, including a novel compound with glucose inserted into the backbone of the molecule. FEBS Lett 393: 273-279.
- López Lara IM, van der Drift KM, van Brussel AA, Haverkamp J, Lugtenberg BJ, et al. (1995) Induction of nodule primordia on Phaseolus and Acacia by lipo-chitin oligosaccharide nodulation signals from broad host range Rhizobium strain GRH2. Plant MolBiol 29: 465-477.
- Broughton WJ, Jabbouri S, Perret X (2000) Keys to symbiotic harmony. J Bacteriol 182: 5641-5652.
- Tariq M, Hameed S, Yasmeen T, Zahid M, Zafar M, et al. (2014) Molecular characterization and identification of plant growth promoting endophytic bacteria isolated from the root nodules of pea (Pisumsativum L.). World J MicrobiolBiotechnol 30: 719-725.
- Saharan BS, Nehra V (2011) Plant growth promoting rhizobacteria: A critical review. Life Sci Med Res LSMR-21.
- Dixon R, Kahn D (2004) Genetic regulation of biological nitrogen fixation. Nature Rev Microbiol 2: 621-631.
- Rees DC, Howard JB (2000) Nitrogenase standing at the crossroads. CurrOpinChemBiol 4: 559-566.
- Teale WD, Paponov IA, Palme K (2006) Auxin in action: Signaling, transport and the control of plant growth and development. Mol Cell Biol 7: 847-859.
- Spaepen S, Vanderleyden J, Remans R (2007) Indole-3-acetic acid in microbial and microorganism plant signaling. FEMS Microbiol Rev 31: 425-448.
- Kawaguchi M, Syono K (1996) The excessive production of indole-3-acetic acid and its significance in studies of the biosynthesis of this regulator of plant growth and development. Plant Cell Physiol 37: 1043-1048.
- Leveau JHJ, Lindow SE (2005) Utilization of the plant hormone indole-3-acetic acid for growth by Pseudomonas putida strain 1290. Appl Environ Microbiol 71: 2365-2371.
- Matsukawa E, Nakagawa Y, IimuraY (2007) Stimulatory effect of indole-3-acetic acid on aerial mycelium formation and antibiotic production in Streptomyces spp. Actinomycetologica. J Soil Sci 21: 32-39.
- Shokri D, Emtiazi G (2010) Indole-3-Acetic Acid (IAA) production in symbiotic and non-symbiotic nitrogen-fixing bacteria and its optimization by taguchi design. CurrMicrobiol 61:217-225.
- Goldstein AH (1986) Bacterial solubilization of mineral phosphates: Historical perspectives and future prospects. Am J AlternAgricult 1: 57-65.
- Hilda R, Fraga R (1999) Phosphate solubilizing bacteria and their role in plant growth promotion. BiotechnolAdv 17: 319-359.
- Fasim F, Ahmed N, Parson R, Gadd GM (2002) Solubilization of zinc salts by a bacterium isolated from air environment of a tannery. FEMS MicrobiolLett 213: 1-6.
- Chen YP, Rekha PD, Arun AB, Shen FT, Lai WA, et al. (2006) Phosphate solubilizing bacteria from subtropical soil and their tricalcium phosphate solubilizing abilities. Appl Soil Ecol 34: 33-41.
- Agrios GN (2005) Plant pathology (5th Edn). Elsevier Academic Press, London, UK.
- Tariq M, Yasmin S, Hafeez FY (2010) biological control of potato black scurf by rhizosphere associated bacteria. Braz J Microbiol 41: 439-451.
- Fargas FRD, O’Hara GW (2006) Isolation and characterization of rhizosphere bacteria with potential for biological control of weeds in vineyards. J ApplMicrobiol 100: 946-954.
- Pechy-Tarr, M Bruck DJ, Maurhofer M, Fischer E, Vogne C, et al. (2008) Molecular analysis of a novel gene cluster encoding an insect toxin in plant-associated strains of Pseudomonas fluorescens. Environ Microbiol 10: 2368-2386.
- Lugtenberg B, Kamilova F (2009) Plant-growth-promoting rhizobacteria. Annu Rev Microbiol 63: 541-556.
- Glick BR, Cheng Z, Czarny J, Duan J (2007) Promotion of plant growth by ACC deaminase-producing soil bacteria. Eur J Plant Pathol 119: 329-339.

Open Access Journals
- Aquaculture & Veterinary Science
- Chemistry & Chemical Sciences
- Clinical Sciences
- Engineering
- General Science
- Genetics & Molecular Biology
- Health Care & Nursing
- Immunology & Microbiology
- Materials Science
- Mathematics & Physics
- Medical Sciences
- Neurology & Psychiatry
- Oncology & Cancer Science
- Pharmaceutical Sciences